
The dandelion does not contain Fibonacci. Fibonacci finds itself through the dandelion.
Stand in a field at midsummer and pick one — the white head, the seed-bearing globe that survived. The pappus, the little parachute each seed wears, will be intact only on a fraction of the heads still standing; the rest will already have given their seeds to the wind. Hold the one that remains. Look closely. The arrangement of the filaments, the curvature of the head, the geometry of the seed positions across the surface — none of this is decorative. Every measurable feature has been selected, across millions of generations, by a single criterion: how far the wind will carry the seeds. The flower head is an aerodynamic instrument.
A nineteenth-century naturalist, given a quiet morning and a fresh dandelion clock, would have drawn it. Not a likeness — a construction. The pencil would have begun with a vertical line down the page, the axis. Then two oval shapes drawn around that axis, one slightly inside the other, like a smaller egg sitting inside a larger one. The naturalist would have labelled them ellipsoid one and ellipsoid two — words from the geometry of solids, the names for the egg-shapes around which everything else would be measured.
Between the two ovals he would have drawn a single sweeping curve, spiralling around the axis as it went, climbing the gap between the inner and outer egg. Ellipsoidal helix. This was the path along which the seeds and their parachutes sat — each seed at a different height on the helix, each one rotated a little further around the axis than the one below it. From every position on this helix, the pencil would lift off into hundreds of fine outward-radiating lines: each line a single bristle of a single pappus, the bristles fanning out into a near-perfect parasol around their seed.
In the margin he would have written the number of pappuses is not as in the real. The detail of the seeds is not drawn. This geometrical construction is affected by approximations. He would have meant: this is what the structure is, allowing for the simplifications a drawing has to make.
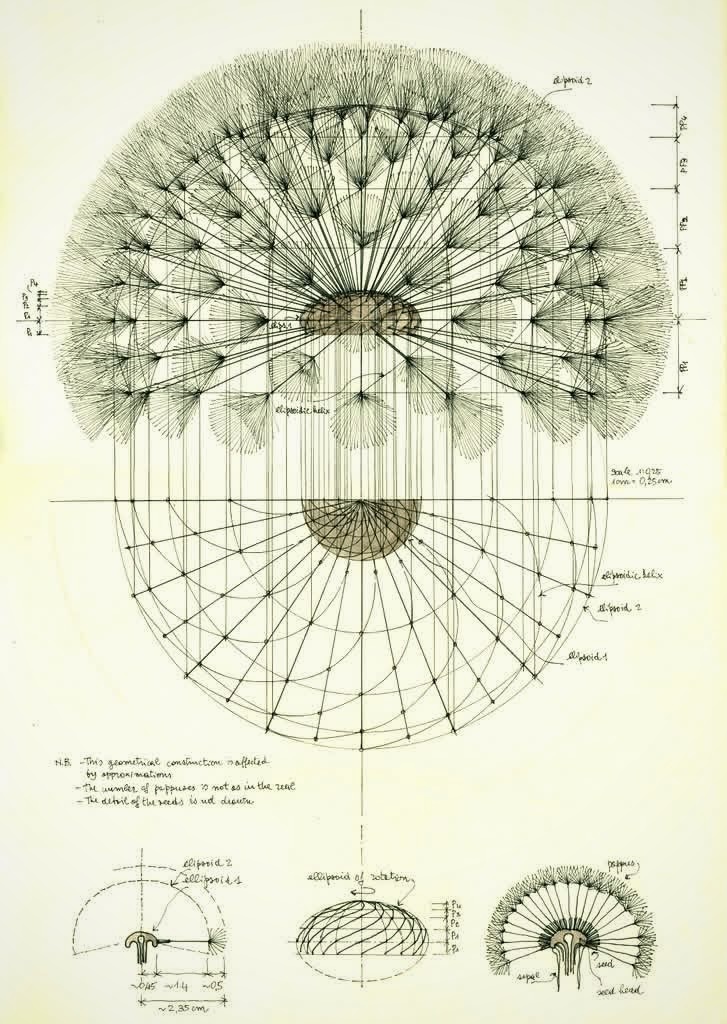
The diagram holds the structure. What follows is what the structure does.
The seed head is not a fluff of more-or-less random fibres around a vaguely round receptacle. It is a structure. Every measurable element — the height of the helix, the angle between one seed and the next, the diameter of each parasol relative to the weight of the seed it carries, the density of the filaments inside each parasol — sits at a value that is locked to all the others. None of it is free to vary independently. The naturalist could see this with his eye and his ruler. He could not see what the structure was doing. That answer would arrive nearly a hundred years after he had finished his diagram, in a wind tunnel at the University of Edinburgh.
Begin with one parasol — one pappus, the parachute around a single seed. There are about a hundred fine bristles in it, arranged in a nearly perfect disc, the bristles slender and slightly curved. If you measured how much of that disc is actually filament and how much is empty space between filaments, you would find that the bristles take up about one-twelfth of the area. The other eleven-twelfths is nothing. The pappus is, by area, ninety-two percent holes.
A parachute that is ninety-two percent holes ought, by ordinary intuition, to be a terrible parachute. The seed should fall straight through it. The air should pass right between the gaps. The way a normal parachute works — air piling up beneath a solid surface, with nowhere to go — cannot work on a structure with this much open area. And yet dandelion seeds are some of the best-travelled small objects in nature. On a still summer day, on a column of warm air rising from a sun-warmed road, they go for kilometres. The mechanism doing this work cannot be the mechanism the intuition reaches for.
In 2018, a research team led by Cathal Cummins at the University of Edinburgh worked out what the mechanism actually is. They put a real dandelion seed in a vertical wind tunnel with a thin sheet of smoke flowing past it, and they watched what the air did.
A small ring of air, like a tiny invisible doughnut, formed just above the pappus and stayed there. Not touching the parachute — floating a short distance above it, suspended in the flow. The air inside this ring was moving in a tight little circulating loop, round and round, separate from the air outside it. The ring did not move down with the seed; it held its position relative to the parachute as the seed descended through the air. The seed was falling inside its own private bubble of low-pressure air, and the bubble was held in place by something to do with the gaps in the parachute.
This is what the gaps are for. As the seed falls, some of the air gets through the gaps in the bristles and emerges on the upper side of the parachute. That air, on the upper side, is now moving in a complicated way — and what it does is curl over and form the floating ring. The ring lowers the air pressure just above the parachute. Lower pressure above, normal pressure below, means more air gets sucked up through the gaps from underneath, which feeds the ring, which lowers the pressure further. The whole thing is self-sustaining. As long as the seed is falling at the right speed, the ring holds.
This little floating ring is what fluid dynamicists call, now, a separated vortex ring — separated because, unlike most vortex rings that appear behind moving objects, this one does not touch the object that creates it. Until the Edinburgh team published their work, nothing quite like it had been described in the textbooks of how things move through air.
The aerodynamic consequence is enormous. The Cummins team measured the drag — the force pulling the seed up against gravity — and compared it to what a solid disc of the same outer size would generate. The pappus generated four times the drag of the solid disc. Four times more lift, from a structure that is mostly nothing. The empty space is not the cost of building the parachute light. The empty space is the parachute.
There is one further finding, and it is what makes the structure feel less like a clever evolutionary accident and more like an engineered fit. Every object moving through a fluid has a speed beyond which the smooth flow around it breaks up. You have watched this happen many times. Smoke from a candle rises in a smooth column for an inch or two, then suddenly breaks into curls and folds. Water from a tap runs in a clear glassy stream until you open the tap a little further and the stream goes turbulent. The change is not gradual. There is a particular speed at which the smoothness gives way.
Fluid dynamicists capture this with a number called the Reynolds number — a single value that tells you how a flow of a given speed past an object of a given size is likely to behave. Below a certain Reynolds number, flows stay organised. Above it, they break up. For the dandelion seed and its floating ring, there is a Reynolds number above which the ring breaks up and the trick stops working. The Edinburgh team measured this threshold. They then asked, of every actual dandelion seed they observed in free flight: at what Reynolds number was it flying?
Every single one was flying below its breakdown threshold. Not just some. All of them. The evolved geometry — the porosity, the filament count, the parachute size relative to the seed weight, all of it — sits at exactly the place where the floating ring stays stable across the entire range of wind speeds the seed will actually encounter as it disperses. There is no margin in the design where the trick fails. There is also no margin where the trick is overengineered. The whole structure is tuned to the edge of the parameter that makes it work.
Return for a moment to the naturalist's diagram. The two nested ovals, the helical curve, the radiating filaments. The naturalist drew, without knowing it, the structure that produces the floating ring. He drew the porosity, the geometry, the helix that arranges the filaments. He could see it was lawful. He could see the values were locked to each other. He could not have known why. The drawing held the structure until the wind tunnel arrived to say what the structure was doing.
The geometry that builds the dandelion's parachute is one thing. The geometry that arranges the seeds across the head of the flower is another. And it is much older, much more widespread, and much more strange.
Look at a sunflower at its peak — late summer, the head fully formed, the seeds packed into the centre. The seeds are not in rows. They are not in any pattern that looks regular at first glance. What you see is a swirl of curves spiralling out from the centre, some going clockwise and some going counter-clockwise. Count the spirals carefully and you find a curious thing: the number of clockwise spirals is almost always 21, 34, 55, or 89, and the number of counter-clockwise spirals is the next number up the same sequence — 34, 55, 89, or 144. These are Fibonacci numbers — the sequence in which each new number is the sum of the previous two: 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144. Pinecones do this. Pineapples do this. Artichokes do this. Daisies do this. The seed head of every member of the Asteraceae family — sunflowers, dandelions, asters — does this.
This was noticed centuries ago. Kepler wrote about it in the early 1600s. What was not understood for most of those centuries was why.
In 1979 a mathematician named Helmut Vogel wrote down a simple recipe that, when followed, produced a pattern indistinguishable from a sunflower head. The recipe was three lines long. To place each new seed, start from the centre. Rotate around the centre by a fixed angle — always the same angle, regardless of which seed you are placing. Move outward by a small amount that depends on which seed in the sequence you are placing. Drop the seed there. That is the whole instruction.
The fixed angle Vogel used was 137.508 degrees.
Two questions follow. Why that exact angle? And how does the plant know to use it?
137.508 degrees has to do with how circles and ratios fit together. If you divide a circle into two arcs in just the right proportions — so that the ratio of the whole circle to the larger arc equals the ratio of the larger arc to the smaller — the smaller arc comes out to about 137.508 degrees. That ratio between arcs is a number called the golden ratio, which has appeared in art and architecture for centuries because of its peculiar visual balance. The angle 137.508 degrees is what you get when you ask: what angle around a circle has this proportion to a full turn?
Imagine you are placing seeds around the head of a sunflower one at a time, rotating by some angle between each placement. If you chose an angle that was a clean fraction of a full turn — say, exactly one-third, or one-fifth — then after three seeds, or five, you would be right back where you started. Every subsequent seed would land on top of an earlier one. The disc would fill along just a few spokes, and the rest of the surface would stay empty.
What you want is an angle that never brings you back to where you started. An angle that, no matter how many seeds you place, never lines up exactly with any earlier seed. That requires an angle whose ratio to a full turn is irrational — not a clean fraction. And among irrational numbers, some are "more irrational" than others, in a precise mathematical sense. The most irrational of all is the golden ratio. It is, in a way mathematicians can prove rigorously, the number that is hardest to approximate with simple fractions. Any other angle will eventually almost-line-up with an earlier seed. The golden angle never does. Each new seed lands in the largest gap available in what is already there. The disc fills uniformly. The packing is as dense as it is possible to make it.
That is the answer to the first question. The angle is what mathematics requires if you want to fill a disc with non-overlapping seeds.
How does the plant know to use it? Plants are not, on any reasonable account, doing geometry.
In 1992, a pair of French physicists — Stéphane Douady and Yves Couder — built an experiment that answered this question by removing the plant from it entirely. They took a flat dish, filled it with silicone oil, and rigged a magnetic field above and around the dish. Then, into the centre of the dish, they began dropping tiny drops of a special liquid — ferrofluid, a fluid that behaves like a tiny magnet. They dropped one every few seconds. The magnetic field made the drops drift slowly outward from the centre toward the rim of the dish. The drops, being magnetic and all pointing the same way, pushed each other apart wherever they got close.
That was the entire setup. Drops added one at a time, in the centre. Drift outward. Mutual repulsion. No instruction about angles, no plant biology, no Fibonacci numbers anywhere in the apparatus.
What appeared in the dish, as more and more drops accumulated, were Fibonacci spirals. The drops, finding their own positions by mutual repulsion, naturally ended up separated from each other by angles that converged on 137.508 degrees. They did this because, in a system where each new drop is added at the centre and pushed away by all the drops already there, the position that minimises crowding is the position rotated 137.508 degrees from the most recent drop. The drops were not solving a geometry problem. They were just being pushed by each other. The geometry came out of the pushing.
This is the answer to the second question. The plant does not know the golden angle. The plant grows by adding new little buds — primordia, the technical term — to the growing tip of its stem. The botanist Wilhelm Hofmeister noticed, back in the 1860s, that each new primordium tends to appear in the largest available gap among the existing ones. It is the same rule as the ferrofluid drops. New thing goes where there is most space. With that one local rule, applied over and over as the plant grows, the golden angle is what falls out. The plant is not performing the calculation. The plant is the calculation, happening in matter, in slow motion, across a growing season.
What just happened is more important than it might look. The dandelion seed head is doing something with a known mathematical optimum — fitting as many seeds as possible into a circle without overlap. It is reaching that optimum by a local rule — each new seed in the largest available gap. No master plan. No central coordinator. No knowledge of the optimum anywhere in the system. And yet the optimum appears, reliably, every time, in every plant in the family.
This pattern — a local rule producing a globally optimal outcome that none of the elements have access to — is the door the dandelion opens.
Consider a soap film. Hold a wire frame bent into any shape — a circle, a rectangle, an irregular twist of wire — and dip it into soapy water. Lift it out. A soap film stretches across the frame. The shape that film takes is, mathematically, the surface of smallest possible area that fills the frame's outline. Not approximately the smallest area. Exactly. The film does not measure anything. It just relaxes into the shape where the surface tension is balanced everywhere — and that shape, it turns out, is the minimum-area surface. The film is the answer to a hard geometry problem, but it solves the problem by not solving it. By just being soap.
In 1970, an engineer at NASA named Alan Schoen was working on minimum-area surfaces by mathematics alone — looking for shapes that have zero average curvature everywhere and repeat themselves in all three directions of space, like a crystal lattice. He found a previously undiscovered surface and named it the gyroid. The gyroid is a shape that is hard to picture. Imagine two perfectly intertwined three-dimensional networks, like a pair of perfectly woven labyrinths sharing the same space without ever touching each other, separated by a single continuous surface that twists through everything. Schoen described it on paper. He did not look for it in nature.
In the years since, the gyroid has been found in nature, repeatedly, in places that have nothing to do with each other.
The wing scales of certain butterflies — the Callophrys hairstreaks, with their iridescent green and blue — owe their colour to a gyroid structure on the surface of each scale, where the wavelength of the spacing happens to scatter blue light back to your eye. The gyroid was not in the butterfly's genes as a "build a gyroid here" instruction. The chitin molecules of the wing scale, packing themselves into the available space under their physical constraints, settled into the gyroid because the gyroid is what those constraints permit at lowest energy.
The same gyroid surface appears in the folds of the endoplasmic reticulum, one of the membranous organelles inside every cell of your body. The lipids of the cell membrane, organising themselves under their own packing constraints, find the gyroid.
The same surface appears in certain synthetic plastics — block copolymers, materials made of two different polymer chains joined together. Mix them at the right proportions, and they spontaneously assemble into the gyroid.
The same surface appears in the inner membranes of mitochondria, the energy-producing organelles in your cells.
Four substrates. Butterfly wing scales, the membranes of the endoplasmic reticulum, synthetic plastics, mitochondrial cristae. None of these systems is in communication with any of the others. They do not share an ancestor. They do not share a developmental program. The molecules involved in each case are entirely different. What they share is the geometry — and the geometry keeps being arrived at because, given the local constraints each substrate is working with, the gyroid is what the constraints allow.
Look at the inside of your own arm. Beneath the skin, the vessels carrying blood divide and divide and divide again — large vessels splitting into smaller ones, those splitting into smaller still, all the way down to capillaries thin enough that red blood cells have to pass through them single file. Look at a tree. Trunk into major limbs, major limbs into smaller branches, smaller branches into twigs, twigs into the veins of each leaf. Look at a satellite image of a river system. One main channel, fed by tributaries, fed by smaller tributaries, fed by streams, fed by trickles.
These are all branching networks. They are doing different things in different materials. But there is a question that applies to all of them. When a parent channel splits into two daughter channels, what should the relative sizes of the daughters be, if the network as a whole is being arranged for least cost?
In 1926, a physiologist named Cecil Murray worked out the answer for a blood vessel. The way he framed it was elegant. A vessel costs the body in two ways. It costs energy to pump blood through it — and that cost goes up steeply as the vessel gets thinner, because thinner vessels resist flow more strongly. It also costs energy to maintain — and that cost is proportional to the volume of blood the vessel holds, which goes up as the vessel gets thicker. Thin vessels are cheap to maintain but expensive to pump through. Thick vessels are cheap to pump through but expensive to maintain. There is a single radius at which these two costs balance most efficiently.
Murray did the calculation. The result was simple. If you take the radius of the parent vessel and cube it, you get a number equal to the sum of the cubes of the radii of the two daughter vessels. That single relationship — Murray's law — describes the diameter ratios in the human arterial tree to a good approximation. It describes the diameter ratios in the bronchi of the lung, where the fluid being moved is air rather than blood. It describes, with remarkable fidelity, the diameter ratios in the xylem of trees — the vessels that carry water from roots to leaves in a plant that has no heart, no pumping circulation, and nothing in common with mammalian biology. It describes the veins of broad leaves. It describes the tunnels of leaf-cutter ant colonies. It describes the tributary networks of natural rivers.
The arterial tree and the river network are not in conversation. They do not share a common ancestor in any meaningful sense. What they share is the optimisation problem — and whatever solves that problem produces the same relationship between branch sizes. Murray's law is not a fact about arteries. It is a fact about branching networks that minimise a sum of flow and maintenance costs. The arteries, the bronchi, the trees, the rivers — each is one substrate in which that optimisation runs to its conclusion.
The third example is older and stranger.
Look at a leopard. The pattern on its hide is not arbitrary. The spots are reliably spaced, reliably sized, reliably distributed. The same is true of the zebra's stripes, the giraffe's patches, the spots on a Dalmatian, the labyrinthine patterns on tropical fish. These patterns appear during the animal's development — they are not painted on, they form in the developing skin as the embryo grows. The question is: how?
In 1952, near the end of his life, Alan Turing — the same mathematician who had cracked Enigma a decade earlier and helped invent the computer — published a paper that proposed a remarkably simple mechanism. Imagine two chemicals diffusing through the developing skin. One of them produces more of itself, and also produces the other. The other inhibits both. The two chemicals diffuse through the tissue at different rates — say, the inhibitor moves faster than the activator. Start with a uniform field where both chemicals are evenly distributed. Wait. The system will not stay uniform. Random tiny fluctuations get amplified by the feedback. Patches of high activator and low inhibitor form, separated by patches where the reverse is true. Eventually the patches stabilise into a pattern. Spots. Stripes. Labyrinths. The specific pattern depends on the rates of diffusion and reaction, and on the shape and size of the field.
Turing was working entirely on paper. He proposed two abstract chemicals he called morphogens. He had no biological evidence. He just showed that, mathematically, this kind of system would spontaneously produce stable patterns from uniform starting conditions.
The chemicals he proposed have since been found, repeatedly, in different animals — different molecules in different species, but doing the same job. The dorsal stripes of certain marine angelfish change with the fish's growth in exactly the way Turing's equations predict on an expanding domain — old stripes spread apart, new stripes appear in the gaps, the spacing relative to the body length staying constant. Photograph a young angelfish, photograph the same individual a year later, and the older photograph has more stripes, interpolated in exactly the places the equations specify.
And then — same mathematics, no biological connection whatsoever — the visual cortex of the brain.
When a mammalian brain develops, the part of the cortex that processes vision organises itself into alternating stripes. Each stripe is dominated by input from one eye; the next stripe over is dominated by input from the other eye. Looked at from above, the visual cortex carries a striped pattern, like a flat zebra hide laid across the back of the brain. The chemicals doing this work are not the chemicals of pigmentation. They are entirely different molecules, in an entirely different tissue, doing an entirely different job. But the mathematics is the same. The reaction-diffusion equations describe both. The cortex is doing what the leopard's hide is doing, and the leopard's hide is doing what the angelfish is doing — not because they have any biological link, but because the mathematics that produces stable stripes from uniform starting conditions is the same wherever it is found.
The fourth example moves from patterns in space to patterns in time. It comes from a desk in a physics lab in the late 1980s.
Imagine a flat table onto which you drop grains of sand, one at a time, at random places on the table. Each grain joins the local pile. Eventually a pile gets too steep, and a little avalanche tumbles down — a handful of grains, redistributing themselves, sometimes triggering small avalanches at neighbouring piles. Drop another grain. Sometimes nothing happens. Sometimes a small slide. Occasionally a big cascade. You keep dropping grains.
In 1987, three physicists at Brookhaven National Lab — Per Bak, Chao Tang, and Kurt Wiesenfeld — built a computer model of exactly this scenario, and found something. The sizes of the avalanches did not cluster around any particular size. There was no "average" avalanche. Instead, the distribution of avalanche sizes followed a particular mathematical pattern called a power law. In plain terms: very small avalanches happened constantly; medium-sized ones less often; really big system-spanning cascades were rare but not vanishingly so. If you plotted the frequency of each size against the size itself, the points fell along a perfectly straight line on a graph where both axes used logarithmic scales. The same pattern across every scale of avalanche.
The deeper point: this happened without anyone tuning the model. No special parameter values were chosen. The model just did this on its own, starting from any initial pile, settling into a state where the avalanche-size distribution always looked the same. Bak and his colleagues called this self-organised criticality — criticality because it is the same kind of statistical signature that physicists see right at the boundary between two phases of matter (the critical point at which water can be either liquid or gas), and self-organised because the system arrives there on its own, with no external help.
The same signature has since been found in earthquake-size distributions, in the sizes of solar flares, in the sizes of forest fires, in the magnetic-domain reversals inside a magnet — and, in 2003, in the brain.
John Beggs and Dietmar Plenz, working at the National Institutes of Health, kept living slices of rat cortex alive on multi-electrode arrays and recorded the cortex's own spontaneous activity. They were not stimulating it; they were just listening. What they saw, when they analysed the recordings, was avalanches — cascades of neural activity rippling across the cortex slice, mostly small, occasionally large, with the size distribution falling on a power law. The same pattern that Bak's sandpile produced. The same pattern that earthquakes produce. The cortex, sitting in a dish, was operating at the same kind of statistical edge as a slowly loaded sandpile or a tectonic fault. The same signature has since been recorded in awake animal brains and in human magnetoencephalography. The brain, by all current measurements, lives at or close to this critical state.
Four areas of nature, four substrates with no shared ancestry, no shared molecules, no shared developmental machinery. Phyllotaxis in plants. Minimal surfaces in butterfly wings and cell membranes and synthetic plastics and mitochondria. Branching networks in arteries and bronchi and trees and rivers. Reaction-diffusion in leopard hides and angelfish and the visual cortex. Self-organised criticality in sandpiles and earthquakes and brains. The biologists and physicists and mathematicians who described these patterns were not, in any meaningful sense, talking to each other. The fields are too far apart. The publication venues too different. The vocabularies too unshared.
And yet every one of these findings has the same logical shape. A local rule, applied iteratively, producing an outcome that is globally optimal — though none of the elements doing the local work has any representation of the global outcome.
So what is the structure that makes this happen?
Think of it this way. None of the elements in any of those examples knows what the system as a whole is doing. The single primordium on the sunflower head does not know it is participating in a Fibonacci spiral. The patch of soap molecules in a film does not know it is solving a minimum-area problem. The single neuron firing in a cortex slice does not know it is part of a power-law avalanche. The single drop of water in a river network does not know it is helping shape a Murray-law branching pattern. And yet each one of these systems, taken as a whole, produces an outcome that mathematicians can prove is optimal — or close to it.
How is this possible?
There is an answer that physics gives, and it has been hiding in plain sight in physics textbooks for nearly three centuries. It goes by a technical name — the principle of least action — but the idea is simpler than the name makes it sound.
Here is the everyday version. Imagine a beam of light passing from air into water — say, looking down at a fish in a pond. The light bends at the surface. The fish is not quite where it appears to be. The bending is consistent enough that it can be measured and predicted precisely.
For a long time, the bending was described by a rule — the angle of incoming light is related to the angle of outgoing light by such-and-such an equation. The rule worked, but it was just a rule, no deeper explanation attached. Then, in the 1600s, the French lawyer and amateur mathematician Pierre de Fermat noticed something. If you ask, of every possible path the light could take between two points — every wiggling, curving, zig-zagging route through air and water — which path is the one the light actually takes? — the answer is: the path that gets the light from one point to the other in the shortest time. Not the shortest distance. The shortest time, accounting for the fact that light travels more slowly in water than in air. The light does not measure distances or times. The light does not compare alternatives. The light just takes the path that minimises travel time. Every time.
Fermat's principle was a glimpse of something much deeper. A century later, the French mathematician Pierre Louis Maupertuis proposed that all of physics works like this. Every motion of every object — a thrown ball, a planet round the sun, a swinging pendulum, a falling apple — is the path that makes a particular mathematical quantity, which he called the action, take its minimum value. Or, more precisely, its stationary value — the value where small changes to the path don't change the action. Maupertuis stated the principle. Euler and then Lagrange, in the eighteenth century, turned it into rigorous mathematics. By the nineteenth century, the Irish mathematician William Rowan Hamilton had extended the framework to electricity, magnetism, and optics.
Then, across the next century and a half, came the recognition that has not been overturned. Every fundamental equation in physics — Newton's laws, the equations of electromagnetism, the equations of quantum mechanics, the equations of general relativity — can be derived from a corresponding least-action principle. The differential equations of physical law and the variational principles of physical law are not separate things. They are the same thing, written two different ways. The local equations describe what each piece of the universe does moment to moment. The variational principle describes what the whole motion, taken as a path, optimises.
This is what the dandelion is doing. The dandelion is not violating the variational structure of physics. The dandelion is one place where that structure is visible at the scale of a thing you can hold in your hand. The seed head is not computing an optimisation problem. The seed head is an optimisation problem solving itself, in matter, by following the local rules its molecules can follow.
It is the same with the soap film. The film is not finding the minimum-area surface. The film is the minimum-area surface — instantly, at every moment of its existence, by virtue of being soap obeying the local laws of surface tension. The mathematics that says "this is the minimum area" and the physics that says "this is what soap does" are not separate facts about the world. They are the same fact, looked at from different angles.
The same goes for the sandpile, the cortex slice, the arterial tree, the river network. None of them is solving an optimisation problem in the way an engineer or an algorithm would. None of them is computing anything. Each of them is, at every moment of its existence, the optimisation it appears to be performing — because the local rules and the global optimum are not two different things.
There is an old phrase that anticipates all of this.
Around the fourth century before the present era — roughly the time of Socrates in Greece — a Chinese text now called the Tao Te Ching recorded a line that has been quoted, studied and pondered in Chinese intellectual life for more than two thousand years. The line is short. Tao sheng yi, yi sheng er, er sheng san, san sheng wan wu. Translated: Tao produces one. One produces two. Two produces three. Three produces the ten thousand things.
The line is worth reading again, without trying to fit it into a religious frame. What the line describes is an iterative generative process. One principle. Step by step, multiplication. At the end: a multitude.
The Chinese phrase 萬物 — wàn wù, the ten thousand things — is the standard way classical Chinese refers to all phenomena, the entire appearing world, plurality named without enumeration. Ten thousand in classical Chinese functions the way myriad used to function in older English — not a count, but an uncountable manyness. So the line is saying, in its plainest reading: the entire appearing world is what one principle produces when it keeps iterating.
This is a description of an attractor system, written twenty-three centuries before mathematicians worked out how attractor systems work.
This series takes its title from this phrase without taking on the metaphysical framework that has grown up around it across the centuries. There is no commitment, here, to whether the Tao is a fundamental principle, a generative grammar, a literary figure, or something else. What the title borrows is narrower. The phrase the ten thousand things captures something real about how the world's multiplicity arises: locally, iteratively, from rules that the products do not contain.
The dandelion is one of the ten thousand things. The sunflower is another. The arterial tree is another. The river network is another. The cortex slice in Beggs and Plenz's dish is another. The reader holding these words is another. Each one is the local running of the same principle, in different material.
What remains is the question. The dandelion does this. The sunflower does this. The cortex does this. Light through varying media does this. The river network does this. They are not in communication. They do not share a developmental history that would explain a common geometric inheritance. What they share is the structure — the variational principle, expressed in whatever local equations the material supports. The question that follows is unavoidable. What kind of universe is structured such that local rules, in every substrate examined, secretly implement global optima?
The question is not rhetorical. It is the question the next five essays will unfold, one substrate at a time, until the substrates are exhausted and only the question remains.
The seed head is small enough to hold in one hand. The structure inside it is, in the most literal sense available to current science, the same structure the variational principles of physics describe. There is no scale at which the two are different. The dandelion does not contain Fibonacci. Fibonacci does not contain the dandelion. Both are local appearances of something that is neither — and that something has been visible in this seed head, on this Tuesday, the whole time you were not looking.